問.答
老師不好意思冒昧請教複製羊的相關問題,
卵細胞來源為黑面,細胞核來源為白面,
不過參考書上寫卵細胞在受精之前就會預先合成胚胎發育初期所需的mRNA,
難道植入細胞核後,原本黑面卵細胞中的mRNA對結果一點影響都沒有嗎?
還是說這些mRNA只在器官發育初期有作用,到後期影響便不怎麼大了?
分化的意義從基因的層次上來說就是部分基因的靜默,即不再表現mRNA的轉錄作用,例如在基因的DNA序列上發生甲基化就能產生此效果。細胞分化是多細胞生物的特徵,不同種類的細胞存在細胞分工現象,因此每種細胞都被關閉了部分的基因,而只表現與功能執行相關的基因群。這種分化過程從胚胎發生的最初階段就開始了,就是你引用參考書的一段話:「卵細胞在受精之前就會預先合成胚胎發育初期所需的mRNA」,這些特定母源基因的mRNA會在卵母細胞內的特定位置累積,而這將預先決定將來胚胎發育時體軸的形成。例如: 果蠅某種mRNA 在卵母細胞後端的累積將決定胚胎時期腹部的形成與生殖細胞的產生。所以植入外來細胞核後,原本黑面卵細胞中的mRNA是會對胚胎發育產生影響的,只不過這些mRNA只在胚胎發育初期有作用,到後期便無影響力了,原因是這些母源基因已經靜默,且mRNA的壽命是很有限的。延伸閱讀:1995 Nobel prize、http://ls.tcu.edu.tw/mdlin/detail.php?recordID=3
嗯......所以卵細胞預先存有的mRNA只是用來決定體軸極性,決定身體發育的大方向,和往後細部的發展(比如說白面或黑面)沒有什麼關係,我這樣的想法對嗎?
再請問一下,生物老師針對卵細胞預先合成mRNA的解釋是:卵裂時速度太快了,來不及轉錄分裂時需要用到的mRNA,所以要先做好;就像這個網站說的:in these early stages the
rate of cell division is so rapid that transcription from the embryonic genome
cannot occur。
有點奇怪,既然分裂得這麼快,快到來不及轉錄,難道就來得及轉譯嗎?還是只是想要簡化問題,用比較簡單的解釋讓我們接受"卵細胞會預先合成一些mRNA"的概念?
我不敢講對或是錯,但我認同你的理解。胚胎發生的最初時期所擁有的某些性狀只具有階段性功能;如同鷹架,用完即可拋棄,而且往後也不再需要,因此預先轉錄在卵細胞內的mRNA,我認為主導的就是鷹架(蛋白)的合成,其他細部性狀的產生就由細胞核內的基因接手控制。
卵裂有一個重要的特徵:細胞分裂之後,細胞越變越小,這與一般細胞必須生長到一定大小才啟動分裂機制有很大的不同,你認為理由何在?此外,卵細胞內含有比例不低卵黃,其中含有大量脂質;內含不少的磷脂與膽固醇(細胞膜的主要成分),又以雞蛋為例,卵黃外還有大量的蛋白。這些事實能否為你釋疑?分裂得這麼快,快到來不及轉錄,難道就來得及轉譯?我想不是來不來得及的問題,而是已經準備好足夠的蛋白質,不須轉譯新蛋白,準備好的卵只是在等待受精後的卵裂行為,因為一切材料準備就緒,所以才能分裂得如此快速。
卵是有極性的,這個極性從前以為只因卵黃分布不均,現在顯然還要加上mRNA的分布不均,很複雜,我不知道是不是有人已經完全理解胚胎最初行為的來龍去脈,一齊期待吧。
老師
影響物種染色體數目的變因是甚麼?
這問題不太好回答。你是複習到哪個單元時碰到這個問題?
就現存生物而言,染色體數目是各種生物的特徵,而且基於遺傳物質的穩定性,各物種的染色體數是不容許有太大的變動的。因此,正常情況下生物是不容許存在影響染色體數目的變因;這是會產生大規模突變的,而突變通常對生物是不利的。
達爾文的演化論推測出一個必然的結論:現存的生物一定具有一個共同的祖先,這個結論現在有許多證據支持它(為何地球上的生物都使用DNA當遺傳物質?)。所以,你提出來的問題,如果用演化的角度來看,就會比較有意義。
你一定注意到了,現在多數生物細胞內的染色體往往兩兩成對;這是有性生殖的結果(精卵必須受精,精卵各自帶有一套相同數目的染色體),但生物一開始只行無性生殖,顯然後來的有性生殖對生物的演化(生存)較具優勢。請你想像一下原始的大海裡,充斥著各種生物的雌雄配子,各自帶著不同數目的染色體,若不同物種但親緣相近的配子碰巧受精並且還能夠生存及繁衍後代,那麼新的物種就誕生了,同時也創造出與原來物種都不相同的染色體數。
有一種物質叫秋水仙素,它可以用來創造三倍體的無仔西瓜,這是一種簡單的影響物種染色體數目的變因。這個事實可以讓你聯想到什麼?為什麼大自然會產生並保留這種秋水仙素以改變物種的染色體數?這不矛盾嗎?你可以合理化這個現象嗎?(請你把這當作訓練提出假設的練習)
既然有可以改變植物染色體的物質(而且變的更好)
那只要找到對於動物無害的那個變因
應該也可以複製此結果吧
改變染色體就是突變,突變通常有害(講幾遍了?為什麼?你自己回想,我說過),好的突變不是不可能,但機會太低。我們無法得知突變的方向,因此無所謂誘導突變往更好的目標。人們在育種過程中,會使用突變技術,但這過程很像亂槍打鳥;完全逢機,有一句成語可以形容:一將功成萬骨枯,這對實驗生物來講是很殘忍的。
現在要把一個會造成突變的因子施加在動物身上,這恐怕會引來保護動物協會的人士的抗議,所以目前科學家如果能用細胞株做類似實驗的話,他們是不會想進行活體實驗的。
至於複製結果,這是當然的,只要是符合科學方法的運作邏輯,在控制變因皆相同的條件下,相同的一個操作變因必然導致相同的結果。不過動植物細胞畢竟是不同的,適用植物的變因未必在動物上行得通。
老師你好,維管束鞘細胞的葉綠餅退化,
但仍保留未折疊的類囊體,這是否和固氮作用中的異型細胞相似呢?
光系統II大多分布在葉綠餅處,而此處退化了,
也就是刪去光系統II,失去裂解水的功能,
但仍保留循環電子傳遞鏈以製造ATP供給暗反應?
在固氮作用中防止水的裂解是怕氧氣破壞,
在C4植物中是否也是希望以此降低氧氣濃度以防止光呼吸,
同時又可製造一些ATP以供使用呢?
異形細胞與維管束鞘細胞的類囊膜都有退化現象,並且還都只保留了能進行循環式磷酸化作用的光系統 I;前者利用它產生的ATP進行固氮作用,後者則是用來補償C4植物在濃縮、使用CO2過程中的能量耗損。這樣的雷同,難怪你會有如此的聯想,我只能說在演化的過程中,有利的性狀就會被保留下來,然後在我們想像不到的時機,因為克服了某種環境的壓力就被放大了出來。
至於這兩群細胞不約而同地都刪除掉光系統 II,就像你說的,避免產生氧氣以保全固氮作用相關的酵素以及減低光呼吸作用的程度。不過科學家們實在不願意相信光呼吸作用只是演化中殘留下來的遺跡,因此一直在找尋更有力的理由來說明光呼吸作用的存在,抄一段課文給你看:We now know that, at least in some cases, photorespiration plays a
protective role in plants. Plants that are impaired in their ability to carry
out photorespiration (due to defective genes) are more susceptible to damage
induced by excess light.
嗯,維基百科表示光呼吸可以消耗過多的ATP和NADPH,在高溫高光強下氣孔關閉使暗反應停滯時尤為重要。只不過為什麼ATP過多時,容易使PSII產出單態氧呢?
現在我們知道光呼吸過程中可以清除掉部分氧氣,顯然過多的氧氣是會毒害細胞的。
那麼為何過多ATP會造成氧自由基的增加?我猜原因之一是呼吸作用被高比值的ATP/ADP抑制了,當減氧的呼吸作用被抑制之後,氧的濃度又在光反應中持續增加,更糟的是此時氣孔還在環境壓力下關閉。最終結果就是氧氣在細胞內累績,並代謝成能毒害細胞的氧自由基型態。
老師為什麼只是因為排列組合不一樣就能對物質性質有這麼大的影響?像是碳能導電而鑽石不能
這個問題很複雜,不過正可顯示化學是門很有趣的學問。
就以你提到的石墨(不要用碳這個字;碳是原子,石墨與鑽石則是碳原子以不同方式組合而成的不同物質)與鑽石的導電差異來說明。物質能導電是因為它的結構中存在可以游離的電子;該電子原本位在離原子核最遠的外層軌域被稱為價電子,若它們沒有與其他原子共用(未被束縛),在特定電場中這個電子就能移動形成電流。石墨的分子結構中每個碳原子都接了三個碳原子;但碳原子有四個價電子,於是留下一個可游離的電子,故石墨可導電。相對的,鑽石分子中的碳都接滿四個碳原子,換言之,每個碳都無價電子可用以游離,故為非導體。
物質的化學特性通常決定於價電子,而原子間與分子間的重新排列組合可以改變價電子的量與性質,因此,排列組合不一樣就能對物質性質產生很大的影響。
熱的物體膨脹是因為分子來回運動把周圍的分子推開,那麼水的冷漲熱縮是怎麼回事?
水在冷卻過程中,水分子間的排列會由混亂狀態變成規律的結晶(冰晶),而低溫結晶狀態的水所佔據的空間比高溫液態時來得更多,因此水在低溫環境下結冰時的體積會增加。附上一張圖給你參考。

水在冰點前後的分子狀態。左圖為固態水的結構;右圖則為液態水。看得出來嗎?左側的固態結晶體,水分子間的距離比較大,所以它的體積也較液態水來得大。造成這種現象的原因是,水分子間存在氫鍵(http://zh.wikipedia.org/wiki/%E6%B0%AB%E9%8D%B5)。由於單一氫鍵的能量並不高,因此在較高溫的環境下,特定氫鍵很容易因為其他水分子本身的動能而遭到破壞(水分子彼此間發生撞擊,此動力大於氫鍵的鍵能),不過隨即可以再與周圍其他水分子建立起新氫鍵。隨著環境溫度的下降,水分子的動能跟著下降到不足以瓦解水分子間的氫鍵,此時的水分子開始被眾多的氫鍵束縛住,不再能夠自由的轉動、移動,只能以振動來表現它們僅存的動能(外在指標就是溫度)。水分子的結構中帶有兩個由氫原子組成的正電端與兩個由氧原子組成的負電端,此現象造成每個水分子最多可以和其他水分子形成四個氫鍵;若是在冰晶中則是以一定的角度形成氫鍵並產生規律的四面體。此冰晶四面體所佔有的空間比起不規律角度形成氫鍵的液態水還要大,所以水在冷卻過程中體積會稍微變大。但這種冷脹熱縮的現象只會在冰點附近的溫度區間中發生,超過攝氏四度以後,水還是冷縮熱脹的。
那既然機會如此渺茫 為何需要擔心基改作物會與近緣野生雜交造成基因汙染呢?? 例如基改玉米與玉米的野生近緣種雜交 這兩種已經算是"不同種"了為何需要擔心呢?
基改作物的內容是這些憂慮的原因。有一種基改作物是插入了蘇力菌毒蛋白基因,它可選擇性毒殺鱗翅目昆蟲的幼蟲;還有一類是插入抗除草劑基因,這可避免作物遭除草劑傷害。由於生物有大量繁殖的潛能,因此就算基因汙染現象的發生機率很低,當乘上生物潛能之後,還是會得出一個不小的數值,事實上野外也的確偵查到基因汙染現象,因而可能導致非害蟲的鱗翅目昆蟲受害又或是超級雜草的產生,所以這種憂慮是可以被理解的。然而我還是贊成基因改良作物的技術繼續下去,那是因為,基改作物除了可以解決糧食問題之外,我還認為這類憂慮有點太過憂慮了。當一個性狀沒有天擇壓力的篩選時(例如除草劑不會出現在野外造成選汰壓力),它很有可能被剔除掉以節約能源;這種現象在細菌上已被證明。所以我才會說這類憂慮是種幽靈。
請問近緣生物之間可以"自然"雜交而產生有效後代嗎? 曾看到考卷上敘述:基改作物種植於田野間,可能會與 近緣植物產生雜交,造成基因汙染
請問不同種植物之間為什麼可以"自然雜交"? 都已經是不同種了
如果真的成功的話,那子代是否是有效後代?
種子植物的自然雜交過程就是授粉,如果授粉的媒介不是特化的動物而是風或是其他沒有選擇性的力量,那麼種子植物其實是非常容易發生不同種的雜交現象,但植物還是有專屬於它們的生殖隔離機制;例如錯開開花時間、柱頭上的養分濃度差異……所以,如果不同種生物產生的有效子代指的是有生殖能力的子代的話,那麼發生的機率將會非常非常低,因為生殖隔離的高牆不容易跨越,但並不是不可能。這個結論適用動物與植物。
老師我想問一個問題,
大家都耳熟能詳脂溶性物質可以用簡單擴散穿過細胞膜,
可是既然它是脂溶性的,為什麼不會就這樣卡在膜上呢?
雖然和濃度梯度有關,只不過這就有點像是萃取,
如果在脂質中溶解度比較好,為什麼還會願意再回到水溶液的界面中呢?
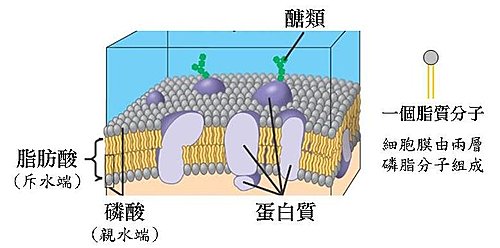
溶解,是以眾多分子間的互動來完成的現象,一個分子能溶解另一個同性質的分子嗎?所以由兩層脂質分子組成的細胞膜對脂溶性物質而言並無溶解的意義。
脂溶性物質可以用簡單擴散穿過細胞膜,其實就是字面上的意思,細胞膜不會去阻擋脂溶性(小分子)物質的擴散,對這些分子而言,細胞膜彷彿不存在似的想進就進、想出就出;擴散作用本就是如此,只要有濃度差,分子們在大的趨勢上就是高濃度流向低濃度。
至於萃取,它是一種分配係數的結果,這世上你大概很難找得到一種絕對親水或是絕對親油的物質,因此萃取需要進行許多次才能提高物質的純度,但我可以肯定的說,純度永遠無法達到100%,意思就是一種物質永遠會同時出現在水與油中。
對,這裡"溶解"一詞應該說是脂質和脂溶性物質"都討厭水",
於是兩者之間相對於水有很好的混合性,可以相容。
所以我很好奇的是,今天脂溶性物質好不容易進到脂膜中,
為什麼還會願意進到"討厭的"水溶液中呢?
例如膜雙層脂質中也是有卡著些脂溶性物質的,像是膽固醇,
那為什麼其他固醇類激素者就會不斷通過,而膽固醇就可以留在膜中呢?
關鍵在於力的平衡。
固醇類激素之所以能進入細胞內是因為細胞內有個受體蛋白能與之結合,這個過程可以提供額外的動力(靜電引力),令固醇類激素擺脫與細胞膜脂質間的疏水性作用力,之後此複合體再送入細胞核啟動基因。(根據BBC的節目指出,細胞質中的骨架絲狀蛋白上存在許多機動蛋白負責將訊息輸往細胞核,我猜前述複合體就是利用此機制,若真是如此的話,這又是另一個動力來源)
如果膽固醇是單純的因為疏水性作用力而卡在細胞膜裡,那麼它是有可能被擠出來的,但是膽固醇在穩定細胞膜上扮演了一定的角色,細胞能容許它輕易地脫離細胞膜?所以合理的推測是,膽固醇與細胞膜脂質間存在非常強的疏水性作用力,又或是兩者間有產生新的化學鍵?這才使得膽固醇能穩定地留在細胞膜中。
老師再次請問,
讀到wobble effect時總有一個疑問:
一組反密碼子
對應 多組密碼子 → 可以成立。
一組密碼子
對應 多組反密碼子 → 正確嗎?
引述華逵選修生物精通:一種密碼子只對應一種tRNA(反密碼子)。
我讀到此時總覺得有點可疑。估算tRNA的種類數目"最小值"約略在32種左右,但生物真正擁有的tRNA種類常比32種多,如此一來不會出現各種tRNA之間密碼子對應的互補?
查了一些資料也沒有直接的答案,大多是反覆重述搖擺效應是"一組反密碼子可對應兩種以上的密碼子"而已,希望老師可以回答,謝謝。
請參閱下列網址:
http://wkl62.blog.163.com/blog/static/50799900201071694853232/
一組密碼子對應 多組反密碼子 → 正確嗎?
從胺基酸與tRNA數量上的對應來看,我想是可以的,前提是特定一組mRNA上的密碼子對應的不同反密碼,其所屬不同tRNA上攜帶的胺基酸是相同的,否則就會產生突變,而突變通常不利於生物存活。所以我們可以做以下的理解:不同生物tRNA數量上的差異,是因為在演化的過程中剔除了那些會導致死亡突變的tRNA。
您好:
我不是您的學生,請恕我冒昧請教您,
馬纓丹花色的遺傳是否純粹顯隱性遺傳?
若生物性狀符合孟德爾顯性遺傳,則其性狀表現有明顯對比;即非黑即白。我查看馬纓丹的花色至少有白、黃、橙、紫、粉紅等表現型,因此不符孟氏顯性遺傳特性。至於馬纓丹花色的遺傳行為屬於哪一類?我猜是複對偶基因遺傳。